
Fos超家族包括c-fos、fosB、fosL1和fosL2基因,这些基因编码的亮氨酸拉链蛋白可以与Jun家族的蛋白二聚,形成转录因子复合物AP-1(Activator Protein-1,AP-1)。AP-1结合 TPA 反应元件和相关的 DNA 元件,如cAMP 反应元件,从而调节一系列生理过程,并在肿瘤的发生、发展中发挥重要作用。
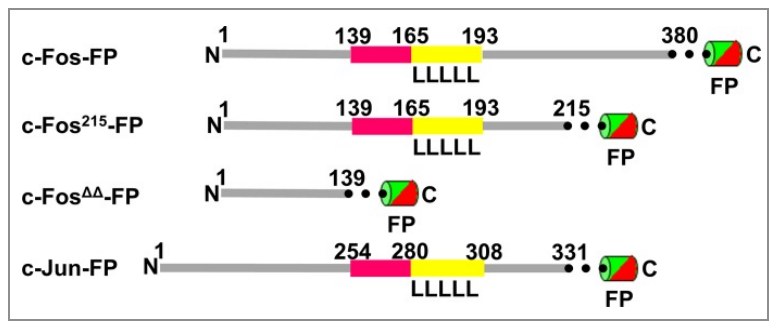
图1 c-Fos结构
c-fos是Fos家族被发现最早和研究多的成员。既往研究发现,c-fos 作为转录因子参与调控细胞的多种病理生理过程,和许多癌症相关。c-fos的mRNA 和蛋白质通常是在刺激后瞬时便开始了转录和翻译,因此被称为即刻早期基因。
c-Fos的细胞内定位和转录活性受到严格调控,特别是转录活性可以通过各种丝氨酸和苏氨酸的磷酸化来增强,包括MAPK p38、ERK1/2,以及ERK1/2活化的激酶Rsk1/2和IκB激酶对它的激活有效。c-Fos蛋白与细胞分化、增殖、存活以及受缺氧影响的组织稳态以及血管生成等关键功能有关;c-Fos也可以对包括胶原酶I在内的多种基因的转录起积极作用。
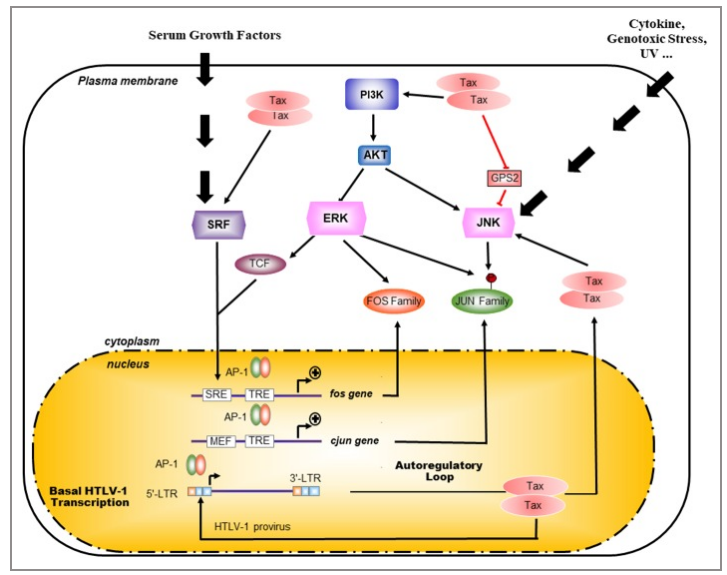
图2 血清等因素刺激fos基因表达
那么,在用WB检测c-Fos蛋白的时候,有哪些需要注意的条件呢?
01
适当地处理激活c-Fos的表达
c-Fos蛋白在某些组织中稳定地表达,但受到不同的刺激时,在许多细胞中被快速瞬时诱导从而高度表达。在后一种情况下,c-Fos的积累在转录、mRNA周转和蛋白质稳定性水平上受到控制。血清、生长因子、肿瘤促进剂、细胞因子、缺氧、营养物质不足和紫外线辐射等均会诱导c-Fos的表达。
a. 血清饥饿处理会抑制c-Fos的表达,添加FBS诱导可以促进c-Fos表达。TPA或者PMA处理激活信号通路。
b. c-Fos易被泛素降解,可尝试MG132处理,以激活蛋白表达和抑制蛋白体降解。
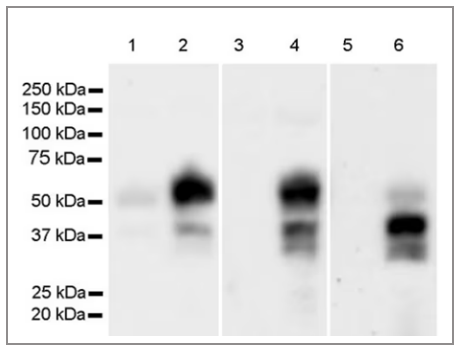
图3 HeLa,NIH/3T3,PC-12,血清饥饿过夜(1,3,5)或PMA/NGF+ MG-132处理(2,4,6)后检测c-Fos
02
根据实验需要,提取核蛋白
当受到特殊处理影响,c-Fos蛋白在细胞质和细胞核的定位不固定。在静止细胞中,c-Fos以极少的量存在于胞质溶胶中。细胞被刺激重新进入生长时,它经历两波表达,第一波在FBS诱导后7.5分钟达到峰值。在这个阶段,蛋白质是定位的内质网。内质网定位需要Tyr-10和Tyr-30的去磷酸化。第二波表达发生在诱导后约20分钟,并在1小时达到峰值。在这个阶段,蛋白质变成核定位。
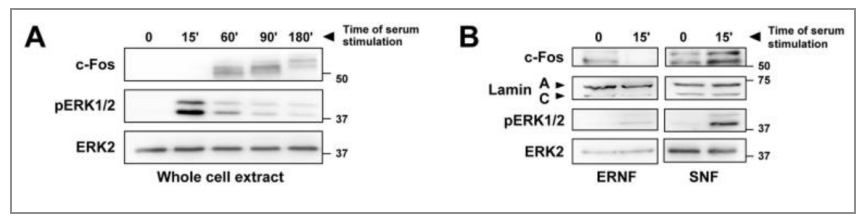
图4 血清刺激NIH-3T3细胞,c-fos在的SNF(可溶性核组分)中积累
03
根据条带趋势分析辨认c-Fos结果
c-fos基因编码62kDa蛋白(380个氨基酸),存在多个剪切异构体,最常见的剪切体分子量41kDa。c-Fos又可与强转录因子c-Jun形成异二聚体,从而形成AP-1复合物。
研究表明,c-Fos蛋白可以在细胞质与细胞核间穿梭。进入细胞核由至少两种核定位信号控制:一种是最有可能利用核导入受体Impβ1的常规基本核定位信号,另一种是位于需要核导入转运蛋白1的蛋白质N端部分的非常规核定位信号。有趣的是,这主要涉及单体c-Fos,并在与Jun蛋白异二聚时受到抑制。这表明二聚化是重要的,不仅对于活性AP-1转录复合物的形成,而且对于将它们保持在细胞核中发挥转录作用也是重要的,c-Fos/c-Jun复合体可影响细胞核的细胞内信号转导。然而,c-Fos的核保留率因其Jun合作伙伴而异,c-Jun比JunB或JunD更强,这与各种c-Fos-Jun二聚体的相互作用强度相关。
从上面几张图我们已经可以发现,使用免疫原序列不同的抗体,即使是相同的细胞中,也可能识别c-Fos不同的异构体。更为特殊的是,某些抗体识别c-Fos与c-Jun结合成的二聚体,它的实测分子量可能远不止41kDa或62kDa。
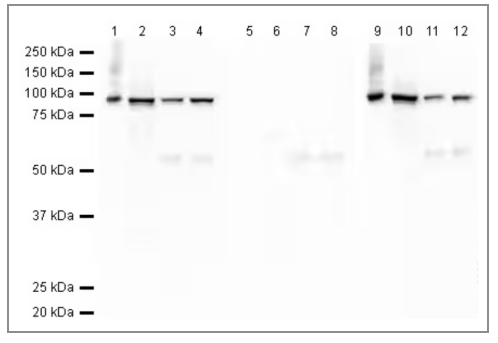
图5 磷酸化c-Fos,二聚体表观分子量约100kDa
来源于优宁维药物研发官网
