
二价基因(bivalent genes),是最初在小鼠胚胎干细胞(mouse embryonic stem cells, mESCs)中发现的一类基因;其启动子区域同时被激活的H3K4me3(组蛋白H3第4位赖氨酸上三甲基化修饰,trimethylation of histone H3 on lysine 4)和抑制的H3K27me3(组蛋白H3第27位赖氨酸上三甲基化修饰,trimethylation of histone H3 on lysine 27)所标记,从而维持转录平衡状态,并且能够在发育过程中被适当的分化信号所激活,促进细胞分化[1]。H3K4me1(组蛋白H3第4位赖氨酸上单甲基化修饰,monomethylation of histone H3 on lysine 4)之前作为预激活增强子(primed enhancer)的标记物被发现也存在于启动子上,并且其在启动子上呈现两种分布模式——与基因激活相关的双峰分布模式以及与基因抑制相关的单峰分布模式[2, 3]。然而,目前的研究仅仅局限于H3K4me1这两种分布模式及其与基因表达之间的相关性,关于H3K4me1在二价启动子上的调控及其在发育过程中的作用在很大程度上还是未知的。
该论文联合生物信息学和实验生物学技术,确定了大部分二价启动子实际上是三价启动子;而这些二价启动子,也就是三价启动子,在发育过程中经历着H3K27me3-H3K4me1转换,通过不同程度的H3K4me1的累积来调控相应组织特异性基因的表达,进而调控组织发育和细胞分化。该研究赋予了H3K4me1一个新的角色,为从表观遗传学角度揭露胚胎早期发育的分子机制提供了重要线索。

作者首先分别根据传统二价修饰(H3K4me3和H3K27me3)以及非传统二价修饰(H3K4me3和H3K4me1)在启动子上的分布模式对启动子CGIs(CpG islands, CGIs)进行分组,发现上述俩种分组结果存在较大程度上的重叠,这意味着大多数传统二价启动子实际上同时被H3K4me1 和二价修饰所标记,暗示着H3K4me1在二价启动子上甚至在发育过程中都扮演重要角色(图1)。

图1 二价启动子同时被H3K4me1和二价修饰所标记
(图源: Yu, Y., et al., Cell Biosci, 2023)
接下来,作者发现在组织发育和细胞分化过程中,这些二价启动子CGIs经历着H3K27me3-H3K4me1 转换:H3K27me3 的丢失伴随着 H3K4me1 的双峰模式丢失或单峰模式富集(图2A)。进一步的研究发现,这种转换可以通过更低程度单峰模式H3K4me1的累积来维持自身组织特异性基因的高表达;同时,通过更高程度单峰模式H3K4me1的累积来抑制其它组织特异性基因的表达,从而促进组织发育和细胞分化,达到谱系建立的目的(图2B)。
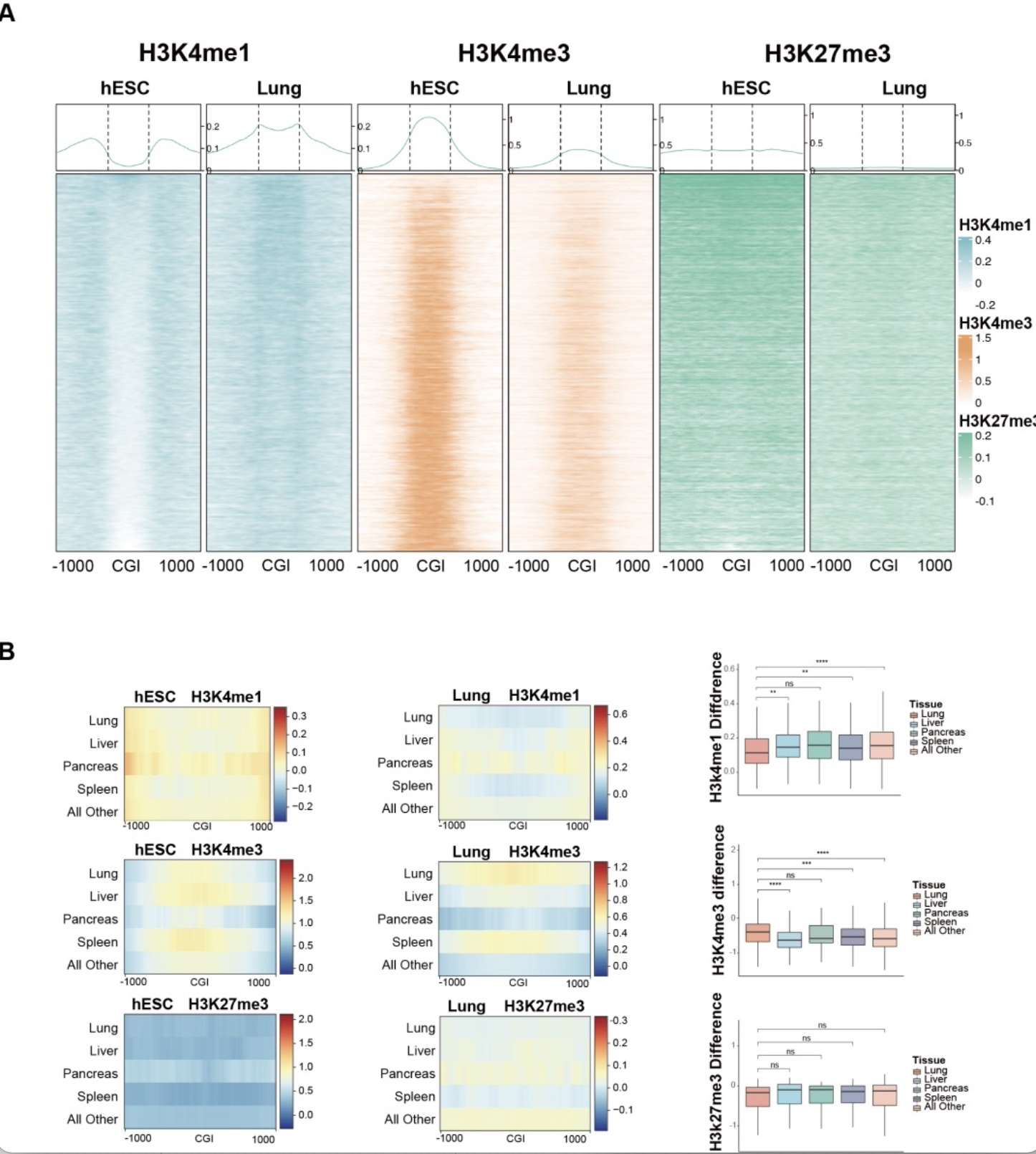
图2 发育过程中二价启动子H3K27me1-H3K4me1转换
(图源: Yu, Y., et al., Cell Biosci, 2023)
同样地,作者在Eed-/-和Suz12-/-mESCs(二者是PRC2核心成员,负责沉积H3K27me3)中发现了artificial H3K27me3-H3K4me1转换(图3A)。进一步的分析发现,该转换可以调控Eed-/-和Suz12-/-mESCs三胚层发育相关基因的表达,表现为:上调内胚层相关基因和下调外胚层相关基因(图3B)。最后,作者利用RA诱导神经外胚层分化实验验证了Eed-/-和Suz12-/-mESCs的神经外胚层分化受损表型(图3C)。
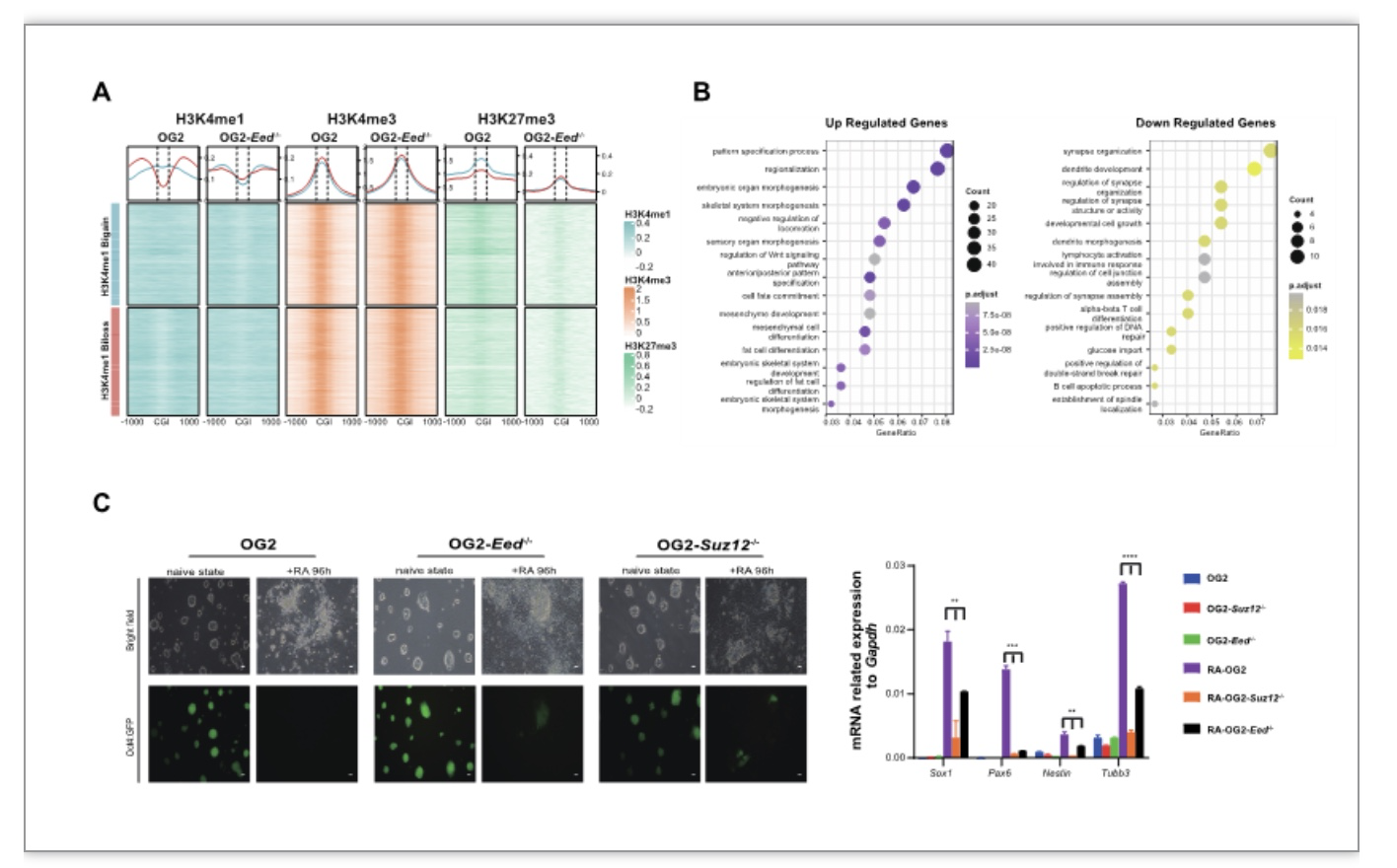
图3 Artificial H3K27me3-H3K4me1转换mESCs调控三胚层分化
(图源: Yu, Y., et al., Cell Biosci, 2023)
进一步,作者利用免疫共沉淀技术、LSD1-ChIP以及CRISPR-Cas9技术,来探究H3K27me3-H3K4me1转换的分子机制。首先,作者发现LSD1与EED和SUZ12存在相互作用(图4A)。其次,LSD1-ChIP结果显示,在Eed敲除之后,LSD1的染色质结合能力下降,导致其不能发挥去H3K4me1甲基化作用,从而建立artificial H3K27me3-H3K4me1转化(图4B)。最后,作者发现,Lsd1-/- mESCs 同样存在artificial H3K27me3-H3K4me1转换,以及外胚层分化受损表型,进一步巩固了LSD1对H3K27me3-H3K4me1转换的媒介作用(图4C-D)。
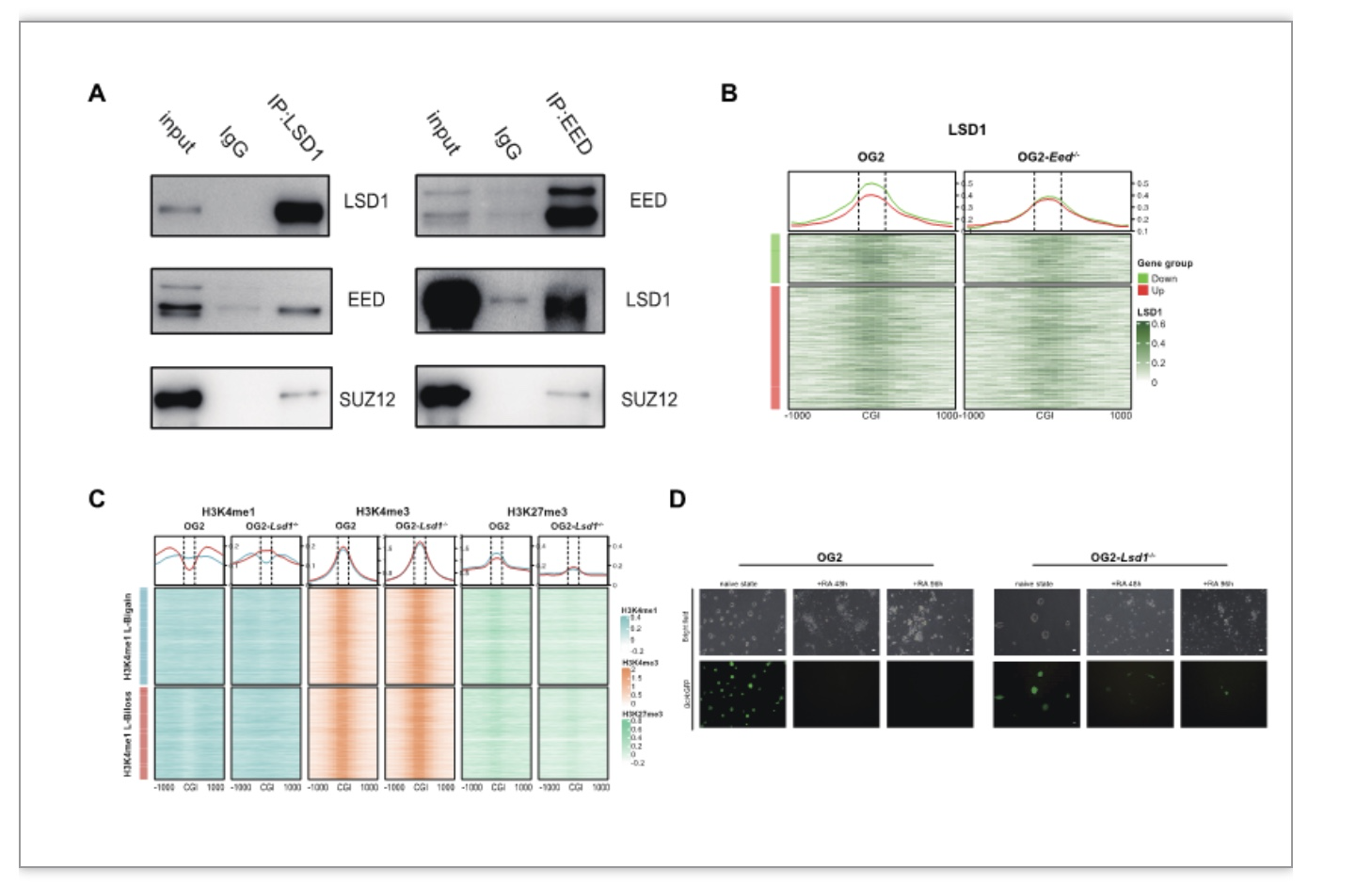
图4 H3K27me3-H3K4me1转换分子机制
(图源: Yu, Y., et al., Cell Biosci, 2023)
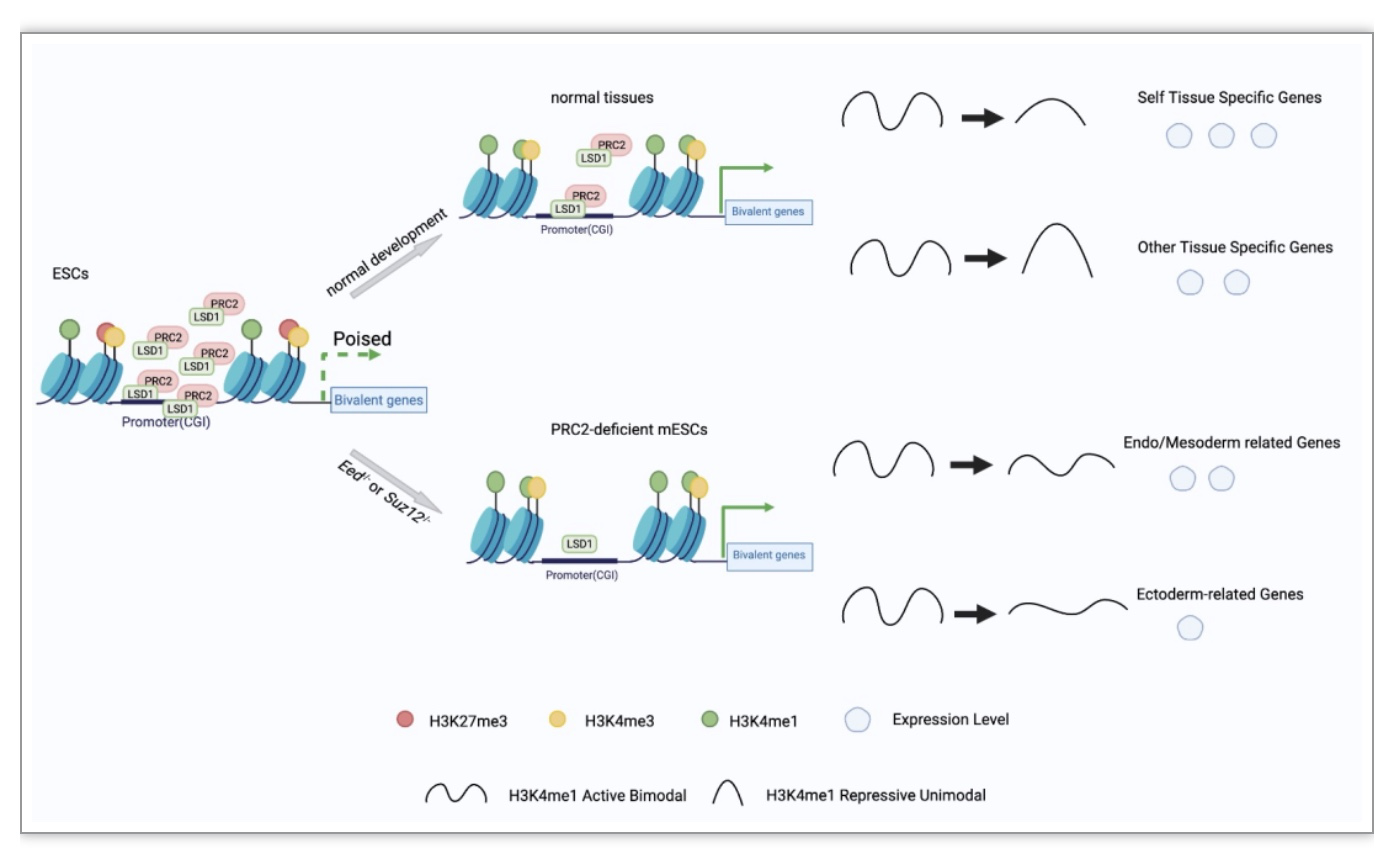
图5 H3K27me3-H3K4me1转换模式图
(图源: Yu, Y., et al., Cell Biosci, 2023)
综上,该研究重新定义了H3K4me1在二价启动子上的作用,作为一个条件性抑制因子,在发育过程中调节组织特异性基因的表达。更重要的是,该研究发现了LSD1-EED/SUZ12通过调节胚胎干细胞中H3K27me3-H3K4me1转换,进而影响早期细胞命运决定;该研究提供了更多的证据从表观遗传角度解释Eed-/-和Suz12-/- mESCs的神经外胚层分化受损表型,从而为后续研究奠定了坚实的理论基础(图5)。
