
基因的表达调控是生物研究的一个重要课题。染色体组蛋白的翻译后修饰和DNA片段的功能密切相关。H4K16乙酰化(H4K16ac)是一个非常特别的组蛋白修饰,它不仅可以调节染色体的凝集程度,而且可以直接影响基因的表达。H4K16乙酰化是由乙酰转移酶MOF介导的。MOF完全敲除的小鼠胚胎发育停滞在囊胚期。这个结果现实MOF和H4K16ac在哺乳动物早期发育中有重要作用。但是MOF和H4K16ac在哺乳动物组织发育中的作用尚不清楚。
作者首先用上皮细胞特异的Krt14-Cre敲除了MOF。免疫组化确认MOF介导的H4K16ac只在皮肤上皮细胞中明显降低而真皮层的H4K16ac不受影响,从而确定了MOF敲除的特异性。上皮细胞敲除MOF的小鼠可以出生,但部分皮肤缺失或者脱落,导致出生后即死亡的表型。对胚胎阶段的研究发现,MOF cKO小鼠皮肤从外胚层到表皮细胞的命运分化是正常的。但随着表皮的生成,Krt14-Cre开始表达,MOF介导的H4K16ac逐渐消失。在胚胎期16.5天(E16.5)的时候,MOF cKO的皮肤明显薄于对照,而且基底膜缺失,细胞极性和增殖遭到破坏,毛囊生长阻滞,细胞凋亡明显增加。

图1 MOF上皮细胞特异敲除小鼠的表型分析
为了避免细胞凋亡带来的间接影响,作者用E15.5的小鼠做了单细胞的转录本分析(scRNAseq)。MOF cKO小鼠的表皮细胞在E15.5的时候已经和正常细胞完全不同了。单细胞的转录本分析显示对照组的细胞分成基底层的祖细胞,上层分化细胞和早期毛囊细胞三个群;而MOF cKO只有两个独立于对照组之外的群,作者称之为类基底层群和类分化细胞群。由于MOF和H4K16ac的作用是直接提高基因表达,作者先分析在敲除细胞里面下调的基因。MOF cKO与对照相比,最明显下调的基因群(Gene Ontology, GO)是细胞核编码的线粒体基因。所以作者认为,MOF介导的H4K16ac能调节线粒体基因的表达。

图2 scRNAseq发现线粒体基因是MOF敲除表皮细胞中最重要的下调基因
因为单细胞的转录本分析只能检测高表达的基因,为了能检测到低表达的基因差异,作者还做了群细胞的转录本分析。在RNAseq中,作者也检测到了线粒体基因的显著下调。但更有意思的是,很多纤毛基因在MOF cKO中也广泛下调。

图3 RNAseq发现在MOF敲除的表皮细胞中大规模的纤毛基因下调表达
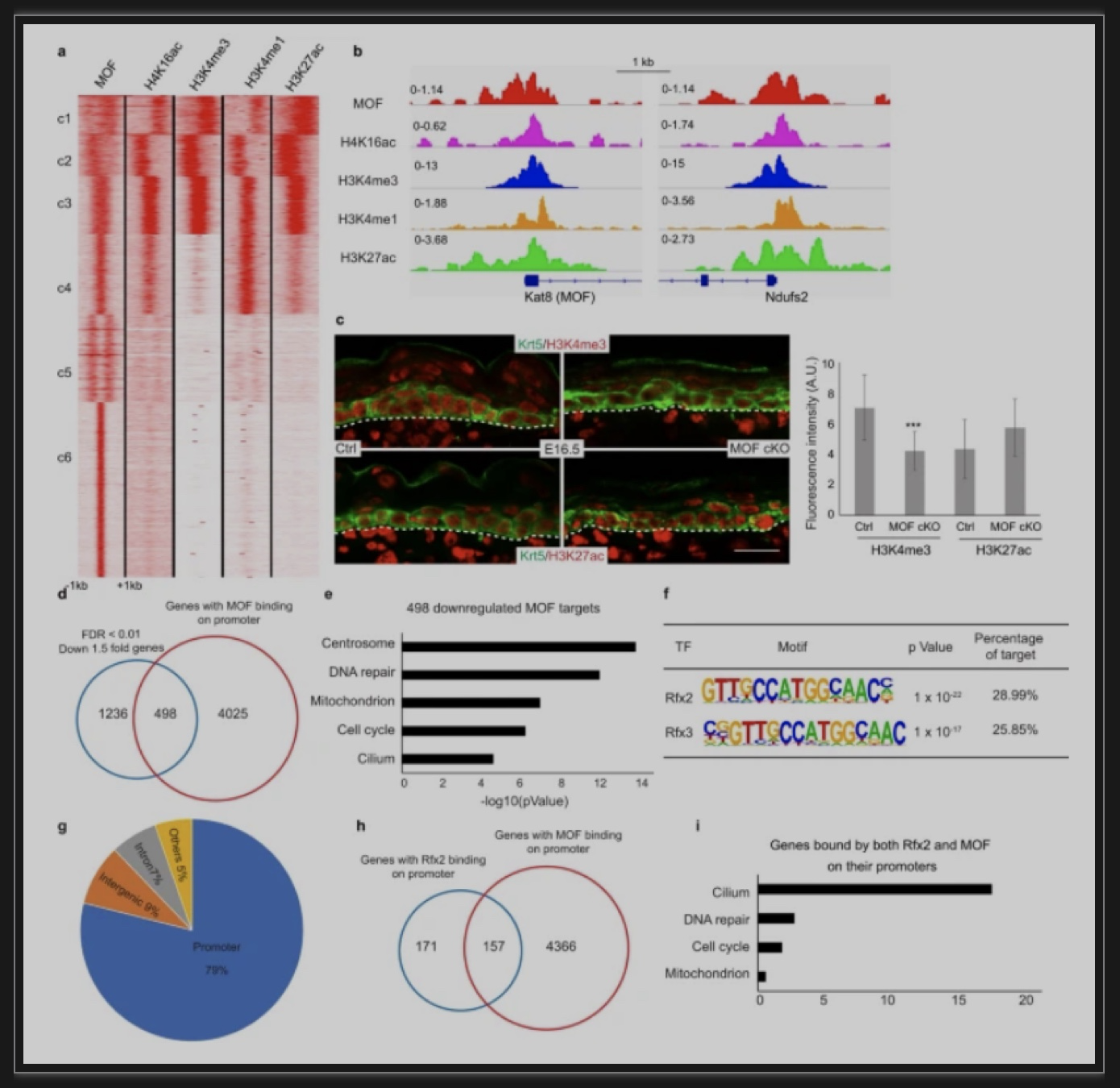
图4 ChIPseq发现MOF主要结合于基因的启动子上
为了研究MOF是如何调节基因表达的,作者做了MOF的染色体沉淀(ChIPseq)。结果发现,MOF主要结合到基因的启动子区域,而且MOF结合的基因很多都是看家(Housekeeping)基因,包括线粒体基因和纤毛基因。这就说明RNAseq和scRNAseq中检测到的线粒体基因和纤毛基因的下调是失去MOF介导的H4K16ac的直接后果。最后,作者发现,一类特殊的转录因子RFX特异结合到纤毛基因的启动子区域调节它们的表达。而在MOF特异敲除的上皮细胞里,这些RFX控制的纤毛基因就不能正常表达。这些结果提供基因调控机理的新线索 - MOF介导的H4K16ac和RFX转录因子在控制启动子为主的转录过程有很强的协同作用。
文章结论与讨论,启发与展望
综上所述,本文作者不仅阐述了MOF及其介导的H4K16ac在皮肤发育中的重要作用,而且深入研究了MOF的作用机制,发现MOF主要是通过结合到启动子来调节基因的表达。MOF调节的基因很多是看家基因,包括两个重要的细胞器,线粒体和纤毛。MOF是如果通过对H4K16ac的修饰调节基因启动子的转录活性的还需要进一步的研究。
