
上一次推送中,向大家简要介绍了蛋白融合表达中需要注意的一些事项,得到了大家的广泛关注和热烈讨论。有很多小伙伴都表示在构建融合表达蛋白的时候出现种种问题,至今还没有得到有效地解决,在这里小编也很无奈,在小编看来,生物学研究属于科学同时也属于玄学,有太多的未解之谜,只能慢慢去尝试解决。

不传播负能量了,来讲点干货:
不少小伙伴都需要在同一个宿主(大肠杆菌或动物细胞等)进行多个外源基因同时表达。此时,有几个不同的策略:
1. 分别将基因构建到多个载体中,共转到同一个宿主,分别进行表达。
这种构建模式是有的,但是经常用于一些特定的应用,比如某些Cas9基因编辑系统,经常采用两个质粒分别进行CAS9蛋白的表达及sgRNA的转录。但是这种模式不推荐用于常规的多蛋白共表达,首先,几个质粒的抗性不应相同,否则容易造成质粒丢失;其次,由于质粒不相容原理,几个质粒不能用相同来源的复制子。介于以上原因,很难构建出稳定的多质粒表达系统。
2. 将多个基因连同各自的启动子放在同一个质粒中,分别进行表达。
这是个不错的解决方案,如果有很高超的载体构建技能的话。很多研究者通过在一个质粒中插入各种复杂的基因元件来实现各种生物学功能,小编我之前就曾在一个质粒中进行三个甚至四个蛋白的共表达,以研究其相互作用关系:
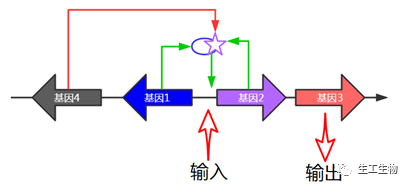
目前市面上有很多这类共表达载体,它们通常携带两个启动子、两个MCS,如老牌公司Novagen的pETDuet™-1:
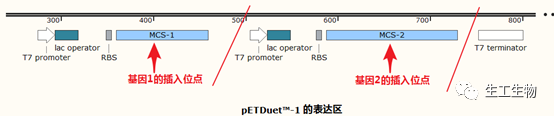
因为在一个载体上放进多个带启动子的基因,通常质粒会变得非常大,增加了构建以及转化的难度。在构建的过程中,要注意两个基因的转录不能形成相互干扰,可以通过添加转录终止位点来缓解不同启动子的转录冲突。另外启动子的选择也是一个不确定因素,如上述商业化质粒采用的是相同的强启动子,在表达过程中,会竞争结合T7 RNA聚合酶,从而造成两个蛋白“此消彼长”的现象。同时,如此多的强表达会对宿主菌带来极大的负担。
3. 多顺反子表达——共表达的理想选择
介于多质粒和多启动子表达的种种局限性,更多的研究者选择多顺反子表达作为解决方案。所谓顺反子,实际上就是基因,单顺反子是指一个启动子启动一个基因的表达,那么多顺反子就是一个启动子启动多个基因的同时表达。
但是并不是说简单地将多个基因排列构建到启动子的后面,就能够实现共表达,在原核及真核宿主中,蛋白的共表达要依赖特殊的功能元件来实现。如果没有这些元件,仅仅能够实现这些基因被转录在同一条mRNA上,通常情况下只有第一个基因能够表达出蛋白。
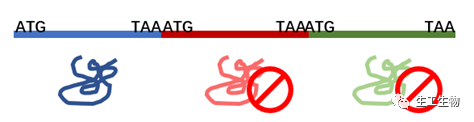
但是别忘了,我们要的是蛋白共表达,与核糖体的结合并起始翻译息息相关,这些元件的作用就是帮助核糖体结合mRNA与翻译蛋白。下面简要介绍一下原核生物和真核生物中可以用到的共表达元件:
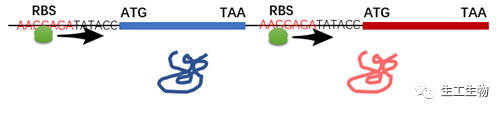
/原核生物:核糖体结合位点RBS/
原核生物的核糖体结合位点(原核中也叫SD序列)位于mRNA的起始密码子AUG上游约8~13nt处,是一段富含嘌呤的4~9个核苷酸组成的共有序列,能与16s核糖体RNA的3端富含嘧啶的区域结合,因此核糖体可以识别、结合这段序列并向下寻找ATG以起始蛋白翻译。在构建多顺反子表达时,务必将SD序列放到基因ATG的前面。获取SD序列的途径多种多样,可以在一些表达载体中找到其序列,如pET-28a的SD序列为AAGGAGATATACC,直接将其构建在基因ATG的前面即可。
由于篇幅限制,先写这么多,关于真核生物的多顺反子表达,请看下篇~

